How the terminal glucoside of the N-glycan donor affects the catalytic efficiency of the eukaryotic oligosaccharyltransferase
How the terminal glucoside of the N-glycan donor affects the catalytic efficiency of the eukaryotic oligosaccharyltransferase
Tropea, B.; Fadda, E.
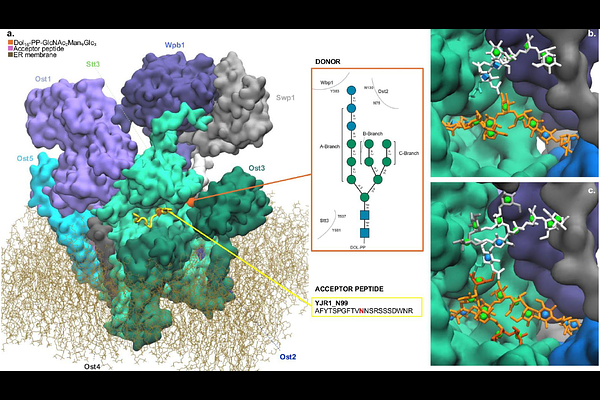
AbstractThe eukaryotic oligosaccharyltransferase (OST) is the enzyme responsible for initiating N-glycosylation of secreted proteins by transferring a pre-assembled lipid-linked oligosaccharide (LLO) donor to target asparagine residues most often found within N-x-S/T consensus sequences, or sequons. OST preferentially selects LLO donors with a distinctive glucoside Glc-(1-2)-Glc-(1-3)-Glc-(1-3)- capping the A-branch. After the N-glycosylation reaction, this motif is cleaved in a stepwise manner from the immature N-glycan structure before the folded glycoprotein exits the endoplasmic reticulum quality control (ERQC) cycle. While the -Glc-(1-3)-Glc-(1-3)- disaccharide is an important flag regulating binding to the calreticulin/calnexin chaperones, the terminal Glc-(1-2)- is removed immediately after OST catalysis, suggesting that its biological function may be directly linked to the OST catalytic efficiency. To understand how and why this capping motif affects the OST N-glycosylation efficiency, we rebuilt 3D models of the yeast OST in complex with an acceptor peptide and LLO donors substrates with and without terminal Glc-(1-2)-, and analysed their stability and dynamics with all-atom molecular dynamics (MD) simulations through both conventional, and Gaussian-accelerated (GaMD) sampling schemes. Our results indicate that the terminal Glc-(1-2)- is essential to anchor the full-length LLO donor to the OST through a complex network of intermolecular contacts extending from the catalytic site to distal subdomains. We show how this contact network is crucial to preserve the LLO catalytically productive alignment of its reducing end. We also show that the removal of the terminal Glc-(1-2)- leads to an increased flexibility of the LLO, which displaces the reducing end and redistributes the conformational ensemble towards misaligned states, which are less catalytically productive. These results provide a mechanistic basis linking the catalytic efficiency of the eukaryotic OST to the distinctive glucosylated structure of the LLO donor.